In 2006, researchers Peter Dodson and Steve Wang estimated that perhaps 71% of all the dinosaur genera that ever existed have yet to be discovered, with majority of the genera that we are likely to find potentially being described within the next 100 years. Whether the estimates are correct or not, there can be little doubt that we are in a “Golden Age of Paleontology” (as far as dinosaurs are concerned, at least), the known diversity of dinosaurs increasing at a prodigious rate. While the majority of the as-yet-unknown dinosaurs are still in the ground, we should not forget that the dusty storage rooms of museums and universities can hold startling fossils, too, as paleontological expeditions often collect more than can be carefully studied and described by the scientists. While not a dinosaur, the discovery of the archosaur Effigia okeeffeae from Ghost Ranch, New Mexico in storage at the American Museum of Natural History, has opened many new lines of inquiry for scientists interested in the Triassic. Not all such forgotten fossils need to represent wholly new groups of animals to be significant, however.
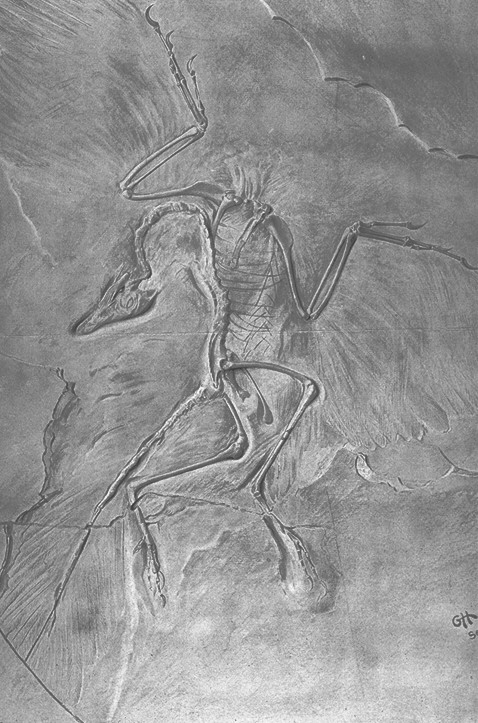
It has often been remarked that if the famous specimens of Archaeopteryx from the lagerstatten of Bavaria did not preserve feather impressions, they would have been deigned small theropod dinosaurs (T.H. Huxley was, as far as I am aware, the first to do this, although I do not have the precise quotation at hand). It isn’t surprising, therefore, that this actually occurred several times, the urvogel turning up again in unexpected places. One of the first to come to light was the Teyler specimen, initially discovered in 1855 (five years prior to the discovery of the single feather described in 1861 by Christian Erich Hermann von Meyer). Labeled Pterodactylus crassipes, the fossil would remain “hidden in plain sight” on display in the Teyler Museum in the Netherlands until John Ostrom correctly identified the fossil in 1970. While possibly only a footnote to the larger story, Ostrom’s discovery created a taxonomy problem as well; because the Teyler specimen was older, traditionally the species name crassipes would have priority over lithographica (Pterodactylus obviously not applying because Archaeopteryx was not a pterodactyl). The name Archaeopteryx lithographica had been used prominently in the literature for over 100 years, however, and so (thankfully) the species name of the early bird remained lithographica.

A replica of the Eichstatt specimen of Archaeopteryx, on display at the AMNH.
After Ostrom’s find, other specimens started to appear, often confused with the dinosaur Compsognathus, also known from the Solnhofen limestone of Germany. In 1973 F.X. Mayr discovered what is now known as the Eichstatt specimen, which he sent to Peter Wellenhofer in order to confirm its true identity. Later, in 1988, Wellenhofer himself discovered another specimen that had been labeled Compsognathus in the collection of the former mayor of Solnhofen, and Wellenhofer again ran into Archaeopteryx in 1992 when a smaller specimen came out of the Solnhofen limestone.
Gerhard Heilmann’s exquisite illustration of the Berlin Archaeopteryx from his work The Origin of Birds.
Such confusion between Compsognathus and Archaeopteryx show the importance of careful examination and taphonomy to paleontology, however; the chief reason why several specimens were misidentified was due to their lack of feather impressions. The exquisite preservation that makes the Berlin specimen of Archaeopteryx a work of natural art is even rarer than the collected remains of the genus itself, and a simple matter of burial environment can seemingly make all the difference. Indeed, in an age where feathered dinosaurs continue to astonish scientists and the public alike, the presence of absence of feathers on larger animals can be problematic. While smaller dinosaurs like Sinosauropteryx and early birds like Confusciusornis are often found preserved in ash falls that allow their discoverers to make out their feather coverings, larger animals may not be covered up as quickly or have such fine detail preserved, as seen from the partial skeleton of Gigantoraptor described in Nature earlier this year. While it is not unreasonable to infer that the giant Oviraptor-like dinosaur had feathers covering its body for at least some of it’s life based upon its relationships to known feathered dinosaurs, no hard evidence of feathers was found, so what sort of feathers it had, how much of its body was covered, and other details remain (for the time being) largely unanswerable. In fact, feather impressions associated with Gigantoraptor may never be found, but some new research involving it’s cousin Velociraptor may provide some clues as to whether the large oviraptorid had plumage or not.
The medium-sized theropod Velociraptor was discovered during the famous American Museum of Natural History expeditions led by Roy Chapman Andrews to the “Flaming Cliffs” of Mongolia during the early 1920’s, and the first remains of Velociraptor to be examined gave the researchers the impression that it was capable of catching relatively large, quick prey with its hands. While certainly an impressive dinosaur, Velociraptor was not as popular as it’s dromeosaur relative Deinonychus, although Gregory S. Paul’s popular book Predatory Dinosaurs of the World started the ball rolling to get Velociraptor to be a household name. While Paul’s book was insightful and prescient in many ways (including its depictions of feathered dinosaurs), the taxonomy in the work was a bit strange, lumping Deinonychus under the genus Velociraptor. This wouldn’t have been of much ultimate consequence, except the book was timed just right to have an important influence on Michael Crichton while we wrote the best-selling novel Jurassic Park, the name Velociraptor being attributed to Deinonychus. This tradition was carried on in the blockbuster film adaptation and in two sequels, the name Velociraptor overshadowing Deinonychus in prestige. As mentioned previously, however, despite the taxonomic reshuffling Paul’s book was important as it drove home the evolutionary relationship between dinosaurs and birds, and in recent years many dinosaurs have come out of Asia showing that they were covered in feathers.
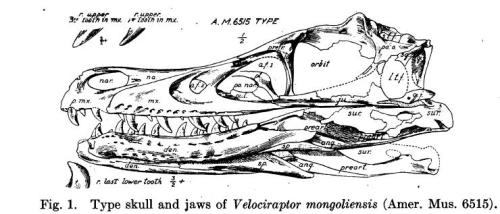
The skull of Velociraptor. From Osborn, H.F., et al. “Three new Theropoda, Protoceratops zone, central Mongolia.” American Museum novitates ; no. 144. 1924
So, how can we tell if dinosaurs that were not find with associated feather impressions had feathers or not? Until now, feathers are often implied for dromeosaurs during at least some stage of life due to evolutionary relationships, but a new (albeit short) paper by Alan Turner, Peter Makovicky, and Mark Norell shows that there are osteological features that tell of the presence of feathers. Along the ulna of a Velociraptor specimen from Mongolia, 14 bumps about 4mm apart were found in a straight line along the bone, directly corresponding to the same structures in living birds, the bumps serving as an anchor for the secondary feathers. This is an amazing find, especially since Velociraptor shows the presence of actual feathers, not just the “fuzz” or integumentary fibers seen on related dinosaurs like Sinosauropteryx. I have to admit that I chuckled a little when I saw one reproduction of Velociraptor covered in feathers, arms obscured by secondaries, but now it seems that such a reconstruction is much closer to the truth than the traditional leathery-skinned model. While the authors of the paper do note that some dinosaurs could have had feathers while the secondary feather anchors were absent, the presence of such a trait gives us a new feature of the bone that can be used to determine whether a dinosaur had feathers or not, and I hope a larger re-investigation of the ulnas of dromeosaurs will be undertaken as it could help determine the presence of feathers on species too big to have them properly preserved.
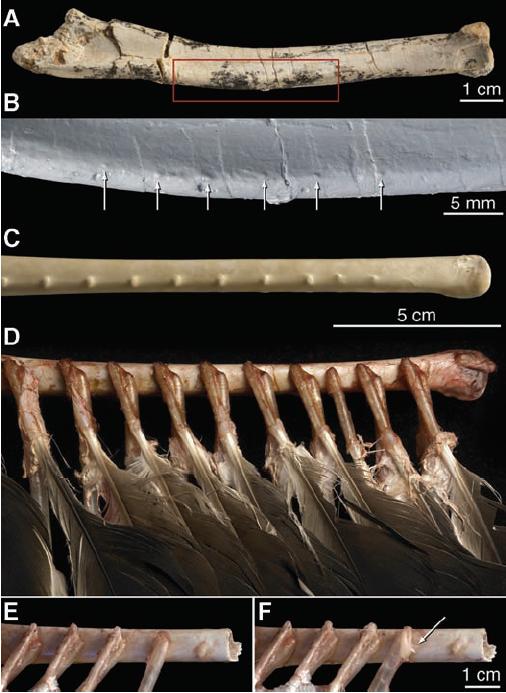
The anchors for the secondary feathers in Velociraptor and a Turkey Vulture. From Turner AH, Makovicky PJ, Norell MA (2007) “Feather Quill Knobs in the Dinosaur Velociraptor.” Science 317(5845):1721.
Still, the question of what was Velociraptor doing with secondary feathers remains. It had previously seemed plausible that many of the non-avian dromeosaurs could have lost some of their feathery coverings, possibly only being covered with feathers as a juvenile. This fossil refutes such a notion for Velociraptor, at least, and secondary feathers could have had any number of uses. While they likely didn’t help much in terms of an individual dinosaur’s thermoregulation, they could have been used for signaling/communication, sexual selection, or been used in the temperature regulation of nests. Personally, I think all these factors could have played a role to a greater or lesser extent, but it is the nest hypothesis that interests me the most.
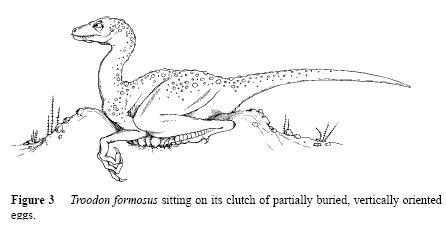
A non-feathered reconstruction of Troodon on a nest. From Horner, J.R. “Dinosaur Reproduction and Parenting.” Annu. Rev. Earth Planet. Sci. 2000. 28:19–45
Those who know their paleo-history will recall that Velociraptor was not the only new theropod to be discovered by Roy Chapman Andrews and his crew. Oviraptor was also uncovered during the expeditions, and the presence of the dinosaur in association with some of the first-known dinosaur eggs gave paleontologists the impression that the theropod was stealing the eggs (hence the name Oviraptor).
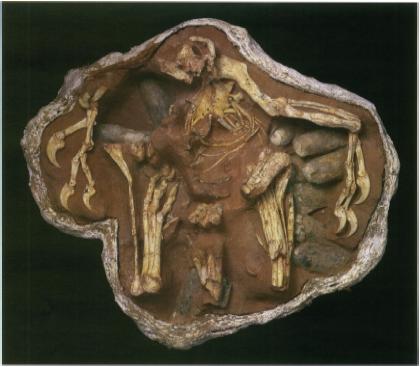
An oviraptorid theropod in a brooding position over a nest. From Clark, J.M., Norell, M.A., and Chiappe, L.M. “An oviraptorid skeleton from the late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avianlike brooding position over an oviraptorid nest.” 1999. American Museum novitates ; no. 3265
Such an interpretation was not to last, however. Research by AMNH staff during the 1990’s showed that the “Protoceratops” eggs that H.F. Osborn and other scientists thought Oviraptor was stealing were really Oviraptor eggs to begin with, the embryo of one of the tiny theropods being preserved inside and allowing for identification of certain eggs with a particular variety of dinosaur. This relationship was further strengthened by the analysis of an oviraptorid dinosaur, probably Oviraptor, in a brooding position on top of a nest. The preservation of this specimen indicates that it died on top of the nest and was not deposited on it after being moved from elsewhere, there being little disturbance to the nest and parent overall. While the discovery of such behavior is momentous in and of itself, if we apply the discovery of secondary feathers in Velociraptor to the oviraptorid (a close evolutionary relative) it would seem that the dinosaur was shielding the eggs with the hypothetical feathers. This is still conjectural, and the oviraptorid would have to be closely investigated to determine whether it had secondary feathers or not, but I don’t think it’s out of the question to infer that, should this oviraptorid be found to have secondary feathers, it was fanning them out over its eggs when it died.

An oviraptorid sitting on a nest, reconstructed as Citipati. From Clark, J.M., Norell, M.A., and Chiappe, L.M. “An oviraptorid skeleton from the late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avianlike brooding position over an oviraptorid nest.” 1999. American Museum novitates ; no. 3265
Given such bird-like behavior in the oviraptorids, it may come as a surprise to find that non-avian theropod dinosaurs may not have had a reproductive cycle like that of modern birds. In a paper released earlier this year, Gregory M. Erickson and others determined that four oviraptorids and one Troodon-like theropod studied seemed to show a more reptilian mode of growth, in that sexual maturity was reached as growth slowed down. This differs from the reproductive modus operandi of living birds, which grow to full size long before breeding begins. While it seems that the dinosaurs, like living crocodiles, took more than a year to reach adult size but attained sexual maturity as adult size was achieved, living birds show explosive growth rates that allow them to reach adult size in much less than a year, yet they are not sexually mature for some time afterwards. Indeed, in dinosaurs it seems sexual maturity was size-linked, while in birds this relationship was decoupled.
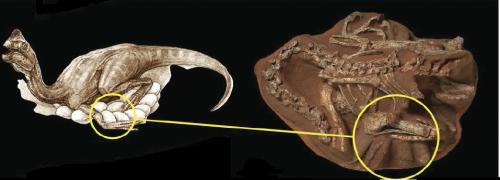
On oviraptorid, Citipati, on top of a nest. From Erickson, G.M. et al. “Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition.” Biology Letters Volume 3, Number 5. October 22, 2007
Despite the difference in growth patterns and life cycles, it is starkly apparent that birds evolved from theropod dinosaurs, some of their closest relatives being the dromeosaurids like Velociraptor. The “big idea” of a evolutionary relationship between dinosaurs and birds has been firmly established, but there are many questions that have yet to be resolved. Helping to further clarify the picture of bird evolution, another recent paper by Alan Turner, et al. (also appearing in Science) describes the new dinosaur Mahakala
omnogovae, which shares a number of features with birds but not later dromeosaurs.

Phylogenetic tree of Paraves, taking temporal factors into account and reflecting changes in body size (click for larger image). From Turner, A.H. et al. “A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight” Science 317, 1378 (2007)
What is surprising about Mahakala is its mix of features and it’s small size. For some time one of the big questions of bird evolution has been “Why did relatively large dinosaurs shrink to take wing?” I had always felt that this was putting the cart before the horse a bit, but now Mahakala has offered up fossil evidence that the large size seen in later dromeosaur celebrities like Velociraptor is a derived condition, the common ancestor probably being no larger than Archaeopteryx.
What does trouble me about this find is it’s age; Mahakala is Campanian (83.5-70 mya) in age. As made clear by the temporal arrangement of the phylogenetic tree, this makes Mahakala much older than Archaeopteryx, Confuciusornis, Yixanornis, and other birds. While Mahakala can tell us much about evolutionary history and has shown that troodontids and dromeosaurids shared a common ancestor which in turn shared a common ancestor with birds (helping to explain those nice secondary feather characteristics in Velociraptor), I am more anxious to see if older, Jurassic relatives can be found. The dinosaurs coming out of Mongolia and China are fantastic finds, but I still find the time disparity between Archaeopteryx and its Cretaceous cousins to be irksome. I’m not the first to bring up such issues either, and I have to say that I do agree with the perspective of Peter Dodson; we need to look at the “big picture” if we’re going to figure this out. In a paper entitled “Origin of Birds: The Final Solution?” Dodson writes;
A philosophy of critical realism seems more congenial for analysis of evolutionary biological individuals having a real history [than cladistics alone]. Cladistics uses parsimony as a first principle, which may be rejected on the grounds that nature is prodigal in every regard. Parsimony based on morphology suffices only when there are no other data sets to consider. Cladistics systematically excludes data from stratigraphy, embryology, ecology, and biogeography that could otherwise be employed to bring maximum evolutionary coherence to biological data. Darwin would have convinced no one if he had been so restrictive in his theory of evolution. The current cladistic analysis of bird origins posits a series of outgroups to birds that postdate the earliest bird by up to 80 million years. This diverts attention from the search for real bird ancestors. A more coherent analysis would concentrate the search for real avian ancestors in the Late Jurassic.
As Dodson notes, morphological analysis alone is not going to get the job done, although I was much relieved by the fact that Turner, et al. used a time scale in constructing their tree. Especially concerning birds, I had always wondered why I would occasionally see animations of Deinonychus growing feathers and flying away as a Canada Goose when Arcaheopteryx was much older. It should be noted that Archaeopteryx is the oldest known bird, not necessarily ancestral to all later birds, but I would hope that more focus would be given to the Jurassic in the search for bird origins as I think the most important fossils to the origins of birds are far older than Mahakala. The chief problem with uncovering the most distant past, however, is that factors of taphonomy might inhibit identification of early bird relatives, especially if they are not preserved in lagerstatten deposits. The fine preservation of so many feathered dinosaurs are partially what has made them so popular, and unless fossil beds resulting from ash falls or ancient lagoons are found, the search for the “early birds” may prove to be exceedingly difficult.
The fossil finds recently reported in Science and elsewhere are definitely important, especially since they shed new light on the evolution of birds and of their dinosaurian relatives. Some, however, have greeted the recent studies with groans; hasn’t everyone had enough of feathered dinosaurs? Such attitudes are unfortunate, as there is still much to learn from specimens that have already been known for a long time. Constant revision and careful reanalysis are the bread-and-butter of good science, and I don’t think any generation of workers should be content with saying “It’s been done” and assume that everything they’ve been told previously is still true. This is not a call to develop new hare-brained hypotheses for their own sake, but rather a plea to keep going back to the dusty shelves of museum basements, to take another look at structures that were initially described decades ago, and to try and keep the bigger evolutionary picture in mind in the search for new specimens. There is too much to learn for any one person to take on these tasks on their own, but as a community I think scientists can still make old bones give up new secrets.
References;
Clark, J.M., Norell, M.A., and Chiappe, L.M. “An oviraptorid skeleton from the late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avianlike brooding position over an oviraptorid nest.” 1999. American Museum novitates ; no. 3265
Dodson, P. “Origin of Birds: The Final Solution?” American Zoologist. Volume 40, Issue 4 (August 2000)
Erickson, G.M. et al. “Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition.” Biology Letters Volume 3, Number 5. October 22, 2007
Horner, J.R. “Dinosaur Reproduction and Parenting.” Annu. Rev. Earth Planet. Sci. 2000. 28:19–45
Nesbitt, S. “The Anatomy of Effigia okeeffeae (Archosauria, Suchia), Theropod-Like Convergence, and the Distribution of Related Taxa.” Bulletin of the American Museum of Natural History. Number 302, Issue 1 (January 2007)
Osborn, H.F., et al. “Three new Theropoda, Protoceratops zone, central Mongolia.” American Museum novitates ; no. 144. 1924
Paul, G.S. Predatory Dinosaurs of the World. Simon & Schuster, NY. 1988
Shipman, Pat. Taking Wing. Touchstone, NY. 1998
Turner AH, Makovicky PJ, Norell MA (2007) “Feather Quill Knobs in the Dinosaur Velociraptor.” Science 317(5845):1721.
Turner, A.H. et al. “A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight” Science 317, 1378 (2007)
Wang, S.C., and Dodson, P. “Estimating the diversity of dinosaurs” PNAS. September 12, 2006, vol. 103 no. 37 13601-13605
















Good summary, but it seemed a bit unfocused. Also, the temporal problem goes away if we consider that perhaps dromaeosaurs are MORE derived than Archaeopteryx (within Aves) but that the Deinonychosauria quickly became secondarily flightless.
And dromaeosaurs are not going to be the answer either way. You can’t posit an avian ancestor from a sister group OR a derived offshoot. I’m convinced that the Paravian precursor is still in Europe somewhere, and NOT in China. All of the attention that Archaeopteryx gets is great–really, the more we learn about the little bugger the better–but the papers musing about its taxonomy and preservation state and whatnot are ignoring the larger issue. Whether Welhoffera is valid or NOT, that still tells us NOTHING about Archaeopteryx’s ancestor.
Archaeopteryx is NOT the first bird (and I get annoyed whenever I see that title), and I await the day when a more primitive bird is discovered. The damn thing is out there–probably in Germany! Somebody find it! 🙂
I thought it was rather good, though I know how hard it is to keep a focussed line of argument going, with dinobirds – everything interconnects. …not that I’m saying it was unfocused 🙂
Anyway, the important thing is that it ended with a healthy hint of cynicism.
The two brooding oviraptorid specimens shown are Citipati, not Oviraptor.
We have many examples of Jurassic dromaeosaurid and troodontid teeth, as well as a partial troodontid skeleton from the Morrison Formation. Then there’s Epidendrosaurus and Pedopenna, paravians from the Middle Jurassic.
The position of Mahakala as a basal dromaeosaurid isn’t very strongly supported. It could easily be a basal paravian or an unenlagiine, for instance.
Finally, that Troodon restoration from Horner’s paper is TERRIBLE. There was absolutely no reason to draw a featherless Troodon in 2000, and I guess that’s why people need to keep getting beat over the head with feathered dinosaur stories.
I noticed in the Mahakala paper that only two features positioned the little critter as a basal dromaeosaur, and I suppose those features could simply be primitive for the group, lost in Avialians and troodontids.
Also, doesn’t Epidendrosaurus’ hatchling status cloud its phylogenetic position?
Not only that, but the authors didn’t code Mahakala for some characters they stated it had/lacked. Currently it falls out as an unenlagiine in my revised version of Senter’s (2007) matrix. I’ll write a report on that after the Rahonavis study I’m doing at the moment.
Epidendrosaurus’ age does hinder understanding of epiphyseal morphology for long bones, but a paravian identity seems universally agreed upon.
You’ll have to send me the Rahonavis treatment, Mickey. It’s one of my favorite “mysterious” maniraptors. Another quick question: In the Mahakala paper, the authors assert that Shanag is closer to the Unenlagiines than to the Micoraptora. Is that accurate? From what I got of the Shanag paper, it’s actually the reverse.
kkkkoooooooooollllllll