Many of the world’s great natural history museums devote at least one hall to creatures that no longer exist today. In the old tradition, in order to keep any young upstarts from getting any ideas about evolution, skeletons or parts of skeletons were grouped by the functions they performed, a visitor being likely to find the wing of a bat and the wing of the bird in the same display case even though the two animals extremely distantly related. Newer layouts, conversely, have largely ignored the end-function of one line or another to group animals together by homology and their shared characters, the most well-known example being the remodeled 4th Floor of the American Museum of Natural History in New York City which has attempted to arrange its fossil collections as a walk-through cladistic diagram.
Still, the generally discarded of grouping animals by their adaptations to general habitats or niches is not without it’s charms. Over and over again, evolution has produced forms that seem to converge on certain body plans, varying habitats making some traits advantageous and others a liability, helping to adapt different organisms to their local ecologies. Flight has independently evolved several times (and the ability to glide an even greater number of times), as well as adaptations to marine environments, saber-like canine teeth, immense sails along the spine, and slicing premolar teeth, although each time such familiar features seem to arise it shows that there is more than one way to solve an evolutionary problem from any given point in an organism’s natural history. Not everything can be chalked up to convergence of form in order to carry out particular functions, however. Parallel evolution, although sometimes difficult to determine, also allows relatively closely related forms to take the same evolutionary paths, showing many of the same anatomical characters even though they diverged from a common ancestor at some point in the past and occupy at least two different lines of descent. In fact, it is often these weird and wonderful creatures that are forgotten or overlooked, more people recognizing the term “saber-toothed cat” (or, loathe as I am to say it, “saber-toothed tiger”) or the genus Smilodon than the term “Nimravid” or the genus Dinictis. The following entry, therefore, will be an attempt to navigate through the somewhat “entangled bank” of evolutionary relationships among animals that appear to be shaped in similar ways by the environment but constrained by their species’ history, showing us that there is more than one way to make a saber-toothed cat.
Back into the pool: Of Ichthyosaurs, Sharks, and Cetaceans
Perhaps one of the most well-known (or at least widely cited) examples of evolutionary convergence has been that of the similar body shapes of sharks, ichthyosaurs, and cetaceans. It’s difficult to see these three distinct groups of creatures side by side and not recognize the similarities, but why are they similar in the first place? If they belong to groups that are distantly-related branches of the evolutionary “bush,” why should they have developed similar body forms?
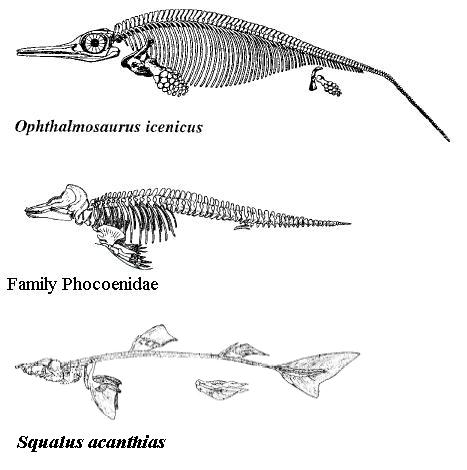
One of the most well-known examples of evolutionary convergence; (From Top to Bottom) An ichthyosaur Ophthalmosaurus icenicus, a Porpoise, and a Spiny Dogfish (Squalus acanthias)
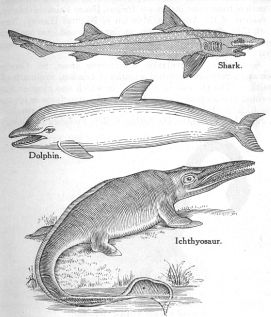
From the 1925 creationist book The Predicament of Evolution by George McReady Price.
Creationists have been quick to seize upon the idea of convergence as if it were one of evolution’s weak points. In 1926, George McCready Price wrote the following in one of the more well-known early American anti-evolution texts, The Predicament of Evolution;
For instance, we have the shark, the ichthyosaur (an extinct kind of fish-shaped reptile), and the dolphin (a true warmblooded mammal, and not a fish at all), all of which greatly resemble each other in external shape and general appearance. Each has the same long, sharp snout, the same powerful tail, the same general fishlike shape. And yet the first of these is a true fish, the second was just as true a reptile, while the third is a mam-mal, bringing forth its young alive and feeding them by milk, just as does a cow or a horse, though it lives in the sea.
Here the evolutionists have to say that this peculiar shape and general form has been evolved separately and independently in each of these three instances. Indeed, Henry Fairfield Osborn, President of the American Museum of Natural History, New York City, declares that a very similar shape and form has been independently evolved “at least twenty-four times.”—”Encyc. Brit.,” Vol. XX, p. 578…
From this large group of facts we become convinced that these many similar or identical structures, which must have been evolved quite independently (if evolved at all), make too great a draft on our credulity. At least, these hundreds of examples of “parallel evolution” greatly weaken our confidence in homology, or similarity of parts and organs, as a proof of blood relationship.
Such arguments have become traditional amongst creationist apologists, suggesting that if convergent evolution does occur then we must throw homology out the window as similar structures will only mislead us as to the true affinities of the creatures being studied. As we will later see with Cuvier’s Ptero-dactyl, this can be a danger for scientists who are unwary and wish to shoehorn creatures into existing taxonomic categories, but not for those who actually look beyond superficial appearances.
The reason why the shark, the ichthyosaur, and the porpoise should all look vaguely the same is because they live(d) in the same environment; the ocean. An organism that is suspended in a fluid that is much denser than air can be adapted in various ways to such an “alien” environment, but physics does dictate what shapes can be taken based upon life history. It is possible to be a floating filter feeder, exhibiting a round shape, but such a strategy is essentially out of the question for animals that need to move quickly and to hunt for food. What is required is not only a powerful propulsive organ to keep the organism moving forward, but also extra appendages to allow for the control of movement and a streamlined shape to reduce drag (and hence reduce energy costs for moving through the water).

One of Charles R. Knight’s renditions of an ichthyosaur.
In fact, sharks as a whole provide a good model for various forms of ichthyosaurs. While ichthyosaurs are generally presented as already being streamlined and possessing a large caudal fin with two equally long lobes, we would be loathe to forget that they too are products of evolution and many fossils show us that they were not always an Euryapsid (thank you, johannes) answer to modern-day Lamnid sharks. Early ichtyhosaurs actually had more of a “bump” towards the back of their tail rather than a full-blown caudal fin, their overall body shape and lack of a large propulsive surface keeping them from moving too quickly through the water. A similar tail type/form can be seen in many modern day sharks like the Nurse Shark, which generally live along the bottom feeding on crustaceans and inhabitants that can be sucked out of coral crevices. Being that ichthyosaurs lack gills, it is unlikely that their early representatives were bottom-dwellers, instead preferring shallow areas, which can be especially productive in terms of food.
Modification of the “tail kink” (which was at first thought to be a taphonomic feature, early reconstructions showing “amphibious” ichthyosaurs with straight tails) seen in early forms allowed for the eventual evolution of a crescent-moon shaped tail, as well as adaptations in the skull and of the limbs into fins (the addition of digits and the addition of bones in the digits being quite common in the latest forms). This more-familiar shape would allow ichthyosaurs maximum propulsion with their caudal fin (the spine going downwards instead of upwards, as in sharks) while they would be able to exert control over their motions with their pectoral fins and would be kept from rolling in the water by their dorsal fins. The evolution of large eyes and other features aside, the overall shape and basic skeletal structure of ichthyosaurs seems to be an optimal design for medium-to-large, fast-moving, oceanic predators (although mosasaurs, pliosaurs, and plesiosaurs took different evolutionary routes).
What allowed ichthyosaurs to develop an effective side-to-side motion of the tail would not work for cetaceans, however. Ichthyosaurs developed their mode of propulsion by side-to-side motions of the spine, perhaps swimming in a mode similar to eels or cat sharks at first, a common form of locomotion in modern reptiles. This sort of motion is usually accomplished on land via a sprawling gait, the limbs being held out to the sides and the animal exhibiting a bit of a side-to-side motion as it moves along.
Whether the ancestors of icthyosaurs were sprawlers (to a greater or lesser extent, predisposing them to side-to-side motions of the tail and body) or not, cetaceans evolved much more recently in evolutionary history, and developed from ancestors that carried their legs directly underneath their body. The plasticity of early archaeocetes and their artiodactyl ancestors was greatly diminished, their hip and spine structure adapted to up-and-down undulations rather than the side-to-side motion seen in the video of the salamander. This sort of constraint has not stopped mammals from becoming adapted to the water, however, and clues to the evolution of cetacean movement can be seen in living animals like Giant River Otters;
In the water, undulations of the spine accompanied with some propulsion from the limbs proves to be very effective, and it’s not hard to imagine an archaeocete like Ambulocetus, as my friend Neil so aptly described, as a “sexy otter.” Once undulation of the spine became established as a method of moving through the water, the eventual addition of a tail fluke would do for cetaceans what the crescent-shaped tail of tuna, sharks, and icthyosaurs acheived in terms of speed and power, the body being adapted towards a streamlined appearance with (again) the pectoral fins providing lift/control and the dorsal fin preventing rolling. Larger forms of whales, namely the Mysticetes or Baleen Whales, grew to immense size and gave up some of the features that seem to be convergent with sharks and the smaller ichthyosaurs (some, in fact, did acheive whale-size), but they are derived from more predatory designs and their niche as massive, far-ranging suspension feeders free them from some constrains while imposing some new ones.

A painting of leaping ichthyosaurs by Heinrich Harder (circa 1916)
Human engineering has recognized similar constraints for motion in the water and even in the air; planes and submarines most closely resemble sharks and dolphins in overall shape, the placement and size of the wings on a 747 having much the same function as the large pectoral fins of far-ranging pelagic fish like the Blue Shark. Life in the water adapted all three groups of animals towards the same shape because there does not seem to be any other way to be a fast-moving, medium-to-large sized marine predator; speed and some degree of maneuverability are paramount. Some other lines have diverged from this shape, as noted before, but the sharks, dolphins, and (I don’t think it’s too much of a stretch to say) ichthyosaurs all occupied essentially the same niche and therefore were adapted in a particular fashion.
Do not think, however, that the convergence of three lines towards one body plan gives credence to a kind of “orthogenesis” or progressive force driving evolution. There was no sort of supernatural or external force manipulating the genetic material of these groups with the shape of a dolphin or shark in mind. Rather, the environment and local ecology determined what form would be favored through time, and even though the three groups may look the same and have significant convergences, they also have many traits in common with their ancestors, allowing us to trace their evolutionary history (which is why no one is arguing that dolphins, sharks, and ichthyosaurs are closely related or form a small monophyletic grouping).
A marsupial you wouldn’t want to meet
Living members of the Carnivora (bears, cats, dogs, civets, weasels, etc.) have always caught my attention, but there was an entire group of carnivorous mammals, now extinct, that have left no living representatives. The last known member of this group was named Thylacoleo carnifex by Richard Owen, and it has some of the strangest dentition ever seen in a marsupial. Marsupial mammals are well-known in Australia, creatures like kangaroos, koalas, and wombats coming most immediately to mind out of living extant taxa. There was a much more diverse population of marsupials during the Pleistocene, however, and the “marsupial lion” was likely a formidable predator.

A skull of Thylacoleo on display at the AMNH.
In order to understand why Thylacoleo is relevant to our discussion of convergence we need to first understand what makes living placental Carnivores so special. Many carnivores, especially cats, have a rather specialized dentition, certain molars and premolars making up what is known as the “carnissal shear.” These teeth are pointed and act like scissors, easily cutting up flesh or crushing bone. The molars behind the shear are often reduced (some groups have retained their molars in order to incorporate a more generalized diet, like dogs and bears), the dental specialization perhaps being one of the keys to the success of this group. Earlier predators of now-extinct lines like Mesonychids lacked such specialized cutting teeth, and the teeth behind the canines of the large Andrewsarchus show that their oral tool-kit was a bit more blunted.

The skull of Andrewsarchus, on display at the AMNH
Thylacoleo, a carnivorous marsupial not descended from the Miacids that gave rise to living carnivores, also developed something of a “carnissal shear” but in a different way. Rather than a battery of teeth that became sharpened, one of the upper and lower premolars of Thylacoleo became elongated and blade-like, and the cleaver-like teeth helped to sharpen each other as they moved past each other when opening or closing the jaw. Thylacoleo also had a terrible bite, the attachments for the muscles that opened and shut the jaw were massive, somewhat constricting the amount of space the brain could take up, but giving Thylacoleo what was perhaps the most powerful bite forces amongst mammalian predators, especially given it’s relatively small body size (it was only about four feet long and 220 pounds).
Thylacoleo is an odd marsupial in another respect; the claw on its thumb was retractable like that of a big cat. This sort of adaptation is especially useful in keeping claws sharp, and perhaps keeping the claws sharp would allow Thylacoleo to get a good hold on its prey before going to work on it with its teeth. At this point I should probably mention that some scholars in the past have thought that Thylacoleo was an herbivore, not unlike the extant marsupial Phalangers. I will leave the response to such an argument to Richard Owen;
These eminent authors received the support, in reference to objections to my conclusions, of the (then) Curator of the Australian Museum, Sydney, Mr. GERARD KREFFT, who, in his contribution to the ‘Annals and Magazine of Natural History,’ series 3, vol. 18, 1866, p. 148, records his opinion that “the famous marsupial Lion was not much more carnivorous than the Phalangers of the present time.”
The species of carnivorous Phalanger is not named. No evidence of such by fossil specimens has reached me, nor have I found such exceptional habit of an existing species of Phalangista elsewhere noted.
As my friend Zach has noted, however, calling Thylacoleo a “marsupial lion” is a bit misleading. Even though some lion-like aspects of the skull (the results of convergence on a hypercarnivorous lifestyle, and Thylacoleo means “pouched lion”) led the anatomist Richard Owen to name the creature on the basis of such resemblances, the ways in which Thylacoleo shows its marsupial affinities are much more important. Referring to this animal as the “marsupial lion” without qualifications (as well as calling the extinct Tasmanian Tiger the “marsupial wolf”) usually confuses more than illuminates, and creationists often take the names and superficial resemblances to mean that evolution didn’t occur. Instead, they propose that God made the beginning of a “kind” of carnivorous mammal which was preserved on Noah’s Ark and gave rise to all later forms, important reproductive habits deemed to be of little consequence.
Even so, Thylacoleo carnifex and its relatives represent a branch of marsupials that became fairly specialized predators, and given the plasticity of tooth structure, it’s not hard to see how sharp premolars could be adapted into a blade to cut flesh. While it may be easy to draw connections between this animal and living carnivores, however, perhaps we should be more measured in our descriptions; both groups met the same challenges in similar ways, but the differences are far more striking and important in this example of convergence on a particular niche.
On what day were the Ptero-Bats created?
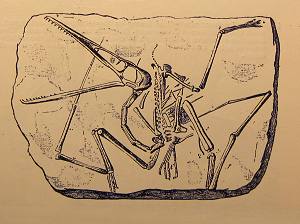
An engraving of the creature now known as Pterodactylus antiquus, the very one described by Collini.
Before there were natural history museums, there were motley assortments of organic odds and ends known as curiosity cabinets, and in the cabinet of Karl Theodor there would eventually come to be a petrified treasure. Although it was probably collected around 1767, the first known pterosaur fossil was not described until 1784, when the appointed caretaker of the collection, Cosimo Alessandro Collini, attempted to determine the nature of the strange creature that came to him from the limestone of Bavaria (the same deposits that later yeilded Archaeopteryx). Although certain that he was the remains of an animal from an earlier time, Collini was agnostic about what kind of animal he had come to possess. Years later, the famed anatomist Georges Cuvier investigated Collini’s paper and illustrations, noting that the creature was certainly a reptile. Still, the fossil would remain without a proper name until Cuvier would write a more detailed analysis in 1809, dubbing the fossil “Ptero-dactyle.”
Not everyone agreed with the analysis of Cuvier, however, especially since Cuvier did not get to see the fossil himself and had to work from the drawings in Collini’s paper. Samuel Thomas von Soemmerring, of the Bavarian Academy of Science, thought that the pterosaur was some unknown type of bat, a view that would remain entrenched in the minds of some scientists for many years. Indeed, one restoration by Edward Newmann in 1843 (and “re-drawn” for Gosse’s work Omphalos, as shown below), depicted the two known types of pterodactyl known at that time as fuzzy bats, complete with cute little ears. It is clear from the drawing that pterosaurs do not make good bats, although this didn’t stop many German paleontologists from taking such a stance through the first half of the 19th century.

Newmann’s “marsupial bats”, conspicuously missing their ears, from Gosse’s Omphalos. It’s likely that Gosse recognized the reptilian nature of these Pterodactyl by the time he wrote his book, so Newmann’s work was copied minus the more mammalian aspects.
But why was there such confusion? It is likely because there is something familiar about pterosaurs that had been seen in living bats; the extension of digits to hold a membraneous wing. While the first fossil, despite wonderful preservation, did not preserve a membrane impression, it is hard to look at it and not recognize the superficially similar structure of a bat’s wing, which also carries a membrane to enable flight. In fact, birds seem a bit unusual in developing feathers for flight; many varieties of gliding and flying creatures have taken to the air (regardless of whether they engage in powered flight or glide) by the use of membranes. Indeed, gliding may often precede powered flight, and once an animal has developed a membrane that can be stretched between its limbs to glide, the extension of the digits at the point(s) of attachment can help to expand the wing size. Such changes likely occur as a result of changes in development, natural selection favoring the invasion of a new niche based upon variations that exist in a population, although in the case of pterosaurs we can no longer test to see if this is correct.
As we just saw with Thylacoleo, however, the convergences of pterosaurs and bats are rather slight, overall. While both acheived flight on membraneous wings attached to extended digits (many more in the case of bats) and have relatively compressed bodies, pterosaurs had a much greater diversity in shape and size than modern bats. Likewise, they did not elongate the rest of their fingers, suggesting that there was some situation (be it climbing or hanging on to a perch) that the pterosaurs still needed their other fingers for (although bats can climb pretty well with their thumbs, and some have even evolved suction disks). Still, it can be said that both took to the air by similar means and had to deal with similar constraints, but their evolutionary paths are far more divergent than that of the aforementioned sharks, ichthyosaurs, and cetaceans.
It doesn’t look like much of a planet-eater to me

A female Gharial at the National Zoo in Washington, D.C.
Perhaps one of the most unnecessarily confusing groups of extinct animals are the phytosaurs. Filling the niche now occupied by reptiles like the Saltwater Crocodile, the water-dwelling archosaurs have left no living descendants despite their past diversity. At first glance, the phytosaur Rutidon looks just like a modern-day Gharial, and even though it shares a common ancestor with the reptiles that now exist in tropical watery habitats all over the world, it is not otherwise related. The most prominent phytosaur feature is that their noses are over or just anterior to their eyes on their head, not at the end of their snouts. This would allow them to breathe while completely submerged, although their eyes might not have been above water when hiding in such a manner. Even beyond this feature, their jaws seem to be fairly simple, merely having a hinge at the back to open-and-close. Compare this arrangement, here represented by the giant Machaeroprosopus gregorii, with the more complex reconstruction of the true crocodilian Deinosuchus (although, admittedly, this reconstruction was heavily based upon the living Cuban Crocodile and may not be fully accurate. It still serves to show the differences between the groups, however).
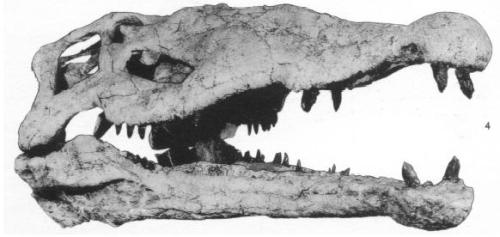
Machaeroprosopus, currently on display at the AMNH

Deinosuchus reconstruction, formerly on display at the AMNH
The most notable difference are the complex bones at the back of the throat of Deinosuchus which are arranged to slide past each other as the jaw opened and closed. No such feature is seen in the giant phytosaur. Still, even after the phytosaurs died out, crocodilians did not return to the water until about the Cretaceous period, many forms being absolutely terrifying land predators that have also long been extinct. One of the early forms was Protosuchus, a small true crocodilian that represented a line that changed little during its tenure on the earth.
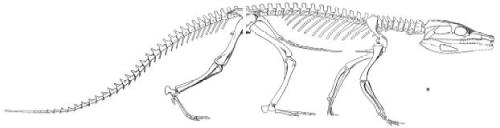
Reconstruction of Protosuchus
Outside of walking relatively high off the ground, Protosuchus had a foreshortened snout which was lower than its eyes, quite different from the arrangement in living crocodilians. As seen in the Dwarf Caiman photograph, below, living crocodilians have their eye sockets on the top of their head, their eyes sticking out on the surface as well as the tip of their nose when they lie in wait for prey (or just rest, for those who would like a less sensationalist tone). Protosuchus, by contrast, has eyes to the sides of the head, even facing somewhat forward, showing that it was much more well-adapted to the land than any swamp or shallow pool. Crocodilans did eventually enter the water, however, and their fossils are among the most common of any vertebrates. Some, like New Jersey’s very own Thoracosaurus, even became marine species, and a few varieties evolved crescent-shaped caudal fins on the ends of their tails to help them swim. The common belief, however, is that crocodiles have always been crocodiles, “changing little since the time of the dinosaurs,” and such generalized half-truths do little justice to crocodilians or their distant phytosaur cousins.

Saber-toothed Nimravid doesn’t sound quite the same…
Many museums have cases devoted to the great saber-toothed cats of epochs long gone, but it would take someone with more than just a cursory understanding of paleontology to sort out what is really being displayed. Saber-teeth, or elongated canines, have evolved many times over in the course of mammalian history, showing up in herbivores like the living Musk Deer as well as extinct groups like the gorgonopsids. Animals as different as a Musk Deer and Inostrancevia are fairly easy to tell apart, even for the non-specialist, but what about nimravids and the “true” saber-toothed cats?
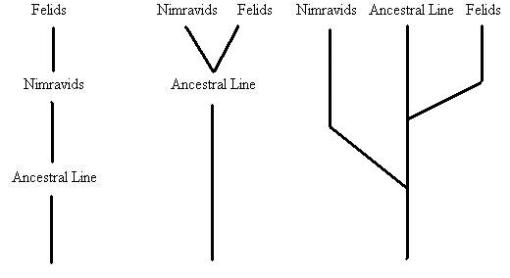
A diagram of the three ideas of Nimravid/Felid evolution.
James Whitcomb Riley is purported to have once written “When I see a bird that walks like a duck and swims like a duck and quacks like a duck, I call that bird a duck.” Unfortunately, this argument is quite popular (even being utilized by the likes of prominent Intelligent Design advocate Michael Behe) despite being very superficial and even vapid. Needless to say, it doesn’t apply to our discussion of Nimravids and true felid saber-toothed cats, but in decades past the two groups were lumped together.
<img src=”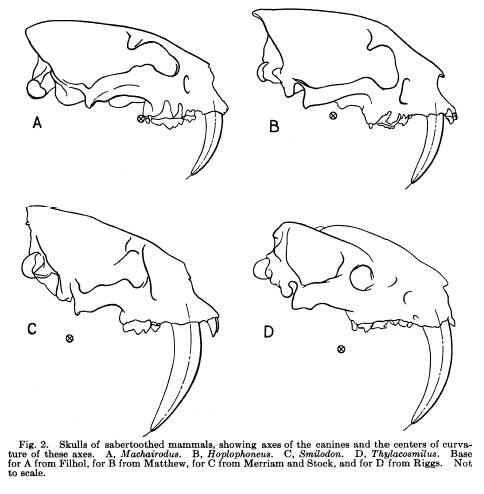 ” alt=”Skulls” />
” alt=”Skulls” />
So, what makes a nimravid a nimravid? They look awfully like cats, so why aren’t they included in the Family Felidae? What makes such distinctions so difficult is that those investigating the skull of Smilodon and Eusmilus would have to be relatively well-versed in scientific jargon and anatomy in order to point out the most important differences. While some nimravids (like Eusmilus) had large canines, their teeth alone are not diagnostic, and the original factors used by E.D. Cope that differentiated these animals from “true” cats were the “alisphenoid canal, postglenoid foramen, carotid, posterior lacerate, and condyloid foramina, postparietal foramina” in the skull (Hunt, 1987). The various canals and foramina listed dictate the paths of various nerves and blood vessels in the skull, and the arrangement in nimravid skulls seem to be more primitive compared with true felids. Likewise, nimravids lack a two-chambered auditory bulla, which is a rounded bit of bone associated with the ear which true cats posess.
There are a few more obvious giveaways when dealing with some nimravids, however. Nimravids equipped with long canines often have more cone-shaped canines than those of saber-toothed cats (which are flatter in cross-section), and many have bony “sheaths” extending from the lower jaw into which the massive teeth fit. Perhaps the most famous example of this kind of arrangement is the genus Barbourofelis, an animal that has actually been assigned to its own family as it is likely more closely related to true cats than nimravids (Barbourofelis was previously classified as a nimravid). Because of this (and the fact that another cat-like offshoot, the marsupial Thylacosmilus) the tooth-sheath shouldn’t be considered diagnostic of nimravids only, but it does give you a substantial clue that you’re probably not dealing with an actual saber-toothed felid.
Despite these differences, it has often been difficult to differentiate the groups (and debate still continues). The diagram above, based upon one in Robert Hunt’s 1987 paper “Evolution of the aeluroid Carnivora. Significance of auditory structure in the nimravid cat Dinictis,” offers three simplified versions of the hypotheses about the relationships of nimravids and felids. Initially it was thought that there was a progressive evolution from ancestor to descendant in a straight line, the nimravids being the direct ancestors to the saber-toothed cats. This view does not represent how evolution truly works, however, and was found to be incorrect. In its place came a view that nimravids and saber-toothed cats diverged from a common ancestor at about the same time, going off in separate directions. This is better, and is more consonant with the data, but again it suggests that the line representing the common ancestor went extinct, either in becoming nimravids or saber-toothed cats. What seems to be the case based upon current data is that the nimravids split off from a common ancestor somewhat before the saber-toothed cats, the line containing their common ancestor continuing its own evolution as both groups evolved. Such a branching pattern is not unusual, and should even be expected, especially since there are living primates like tarsiers and lemurs that represent the overall kind of animal our ancestors once were, but still quite different and undergoing their own evolution alongside our own lineage.

The skull of Thylacosmilus, the marsupial answer to the saber-toothed cat, on display at the American Museum of Natural History in New York City. Note how far back in the skull the roots of its massive canines extend.

The skull of Megantereon on display at the AMNH. It was one of the “true” saber-toothed cats.
To complicate things even further, the skull or skeleton of the marsupial Thylacosmilus is also often thrown into the mix. Although totally unrelated to nimravids or felid saber-toothed cats, the South-American Thylacosmilus converges closely on the appearance of the placental predators, although there are some important differences. As can be seen from the above photographs, the eye of Thylacosmilus is entirely enclosed by bone on the side of the head, while in many felids and nimravids the eye socket is not entirely ringed-in by bone as if someone had bored a hole in the skull (compare the skull of Thylacosmilus with that of Thylacoleo, above). Further, the teeth of Thylacosmilus have very deep roots, going back in the skull almost over the eye. Originally it was thought that the teeth faced outward, but this was based upon a distorted skull and later finds showed the true position of the long canines.
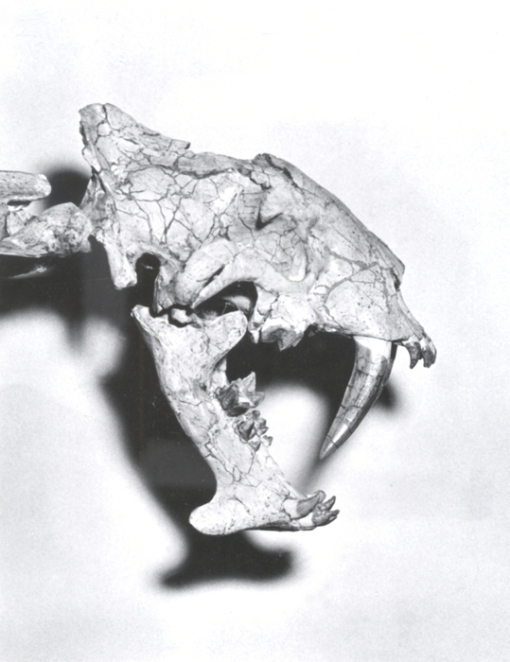
Hoplophoneus
Now that we have elimated Thylacosmilus from the running as another case of marsupial convergence, we must ask why the nimravids and felid saber-toothed cats are so close to each other in appearance. While many of the instances I’ve discussed previously have been instances of convergence, be it throughout the entire body or merely certain aspects of it, the nimravid-felid connection is a wonerful example of parallel evolution. W.E. le Gros Clark provides an excellent summation of understanding the difference in his 1959 book The Antecedents of Man;
From what has already been said, it is clear that, in assessing degrees of phylogenetic affinity, it is always necessary to take into account the factors of parallelism and convergence in the evolutionary development of related or unrelated groups. These processes can lead to structural similarities, which, taken by themselves, may be misleading. The term convergence is applied to the occasion in general proportions or in the development of analogous adaptations in response to similar functional needs. But such similarities are superficial and easily distinguishable by a detailed comparative study of the animal as a whole. For example, the resemblance in general appearance, even in a number of morphological features, of the Tasmanian wolf to a dog does not obscure the fact that in fundamental details of their anatomical construction they belong to quite different mammalian groups. On the other hand, the potentialities of parallelism seem often to have been much overestimated by some anatomists, for this phenomenon has sometimes been invoked in support of extreme claims for independant evolution of groups which are almost certainly quite closely related. We can agree with G.G. Simpson that the whole basis of parallelism depends on an initial similarity of structure and the inheritance of a common potentiality for reproducing homologous mutations, and that, this being so, the initial similarity and the homology of mutations themselves imply an evolutionary relationship. Expressed in another way, it may be said that convergence increases resemblances (which are, however, no more than superficial), while parallelism does not so much increase resemblances as maintain and perpetuate (by development ‘in parallel’ so to speak) similarities which have already existed ab initio in the genetic make-up of related types. Thus, ‘closeness of parallelism tends to be proportional to closeness of affinity.’
There are a few problems with this reasoning, namely that it seems to give credence to an almost pre-determined genetic course for the lines to evolve in parallel, although le Gros Clark makes it clear in the work that he does not support in any way the notion of orthogenesis. Still, the passage makes the important distinction that in order to undergo parallel evolution groups need to be somewhat closely related and already bear similar structures, evolution preserving many of the similar traits instead of working to the same end from two disparate points. In the case of the nimravids and the felids it seems that they evolved from a common ancestor which was probably taken to carnivory. Nimravids branched off earlier, being more “primitive,” while the felids came off the same line (or a very similar one) after it had accumulated a few more evolutionary changes. Indeed, even if form seems to be static or change little, it’s hard for me to believe that designs are not slightly adapted this way or that as if the creature was an already perfect creation not influenced by changing ecological circumstances. Still, it seems that the nimravids and felids were adapted in similar ways, their ancestral lines probably possessing at least semi-retractable claws, long and sharp canines (although not long to the extreme like its descendants), a shortened face, and a developed carnissal shear. It is really not that difficult to change a civet-like creature (or in the case of our hypothetical common ancestor, a creature a bit closer to a cat) into a saber-toothed Smilodon, the changes being modifications of existing structures more than the creation of something entirely new out of nowhere. In fact, the vertebrate tetrapod skeleton has proven to be quite versatile, and most of the major bones in any vertebrate skeleton can be found to correspond with those in another vertebrate, allowing us to compare rhinos with ceratopsians, dromeosaurs with birds, cats with dogs, ichthyosaurs with cetaceans, and humans with primates.
Of constraints and convergence
I hope that is has become clear why convergence is such a strong theme in the evolution of vertebrates. At this point in the history of evolution, vertebrates have had a chance to fill nearly every niche imaginable in a large variety of habitats over millions of years, and so common themes are bound to arise. When groups return to the ocean, the environmental constraints shape them in ways peculiar to their new way of life that would not be advantageous in other situtations (i.e. being such a large aquatic animal that you’d be crushed by your own weight if you came onto land). When mammals become adapted to be predators, their dentition and morphology must be altered if they are to be successful hunters, carnivores past and present showing some suprising similarities despite being only distantly related. Even when taking to the air, laws of physics still apply, and natural selection often works through physical and chemical constraints to produce new forms.
It is of little doubt that the tetrapod design is a versatile one, retaining its overall character through the various changes that it has endured. Indeed, even when a lineage dies out and may seem gone forever, there is no law that says a similar situtation in the future will not produce forms that may be strikingly familiar, even if such organisms are not directly related to the last group that filled their new niche. Evolution has produced “endless forms most beautiful and most wonderful” and will continue to do so long after I am gone, but random mutation/natural selection do not work in isolation from the rest of the natural world. Evolution has produced so many amazing creatures precisely because ecology, physics, and chemistry have offered up both opportunities and challenges, and I only regret that I will not be able to witness the familiar and unfamiliar about what is swimming in the seas, flying in the air, or stalking the land 500 million years from now.











































{kind=link}
{kind=link}
Recent Comments